

Before a discussion of the use of faunal resources by the inhabitants of the Wall and Fredricks sites can be attempted, the state of preservation of the two faunal assemblages should be evaluated. As noted earlier, the contexts from which the bones were retrieved at the two sites were dissimilar. A majority of the bones from the Fredricks site were recovered from burial pit fill, whereas a majority of those from the Wall site were recovered from deposits of sheet midden. It has been suggested that "small fragments just would not survive" in a midden deposit (Runquist 1979:342) and that bones deposited in pits are less likely to be stepped on, exposed to scavengers, or damaged by weather than are bones which are not placed in pits (Chaplin 1971:16; Waselkov 1977:84).
At the Wall site, 19.96% of the bone was retrieved from 1/2-inch screen, 65.07% from 1/4-inch screen, and 14.97% from 1/16-inch screen. At the Fredricks site, 20.91% was recovered from 1/2-inch screen, 70.12% from 1/4-inch screen, and 8.96% from 1/16-inch screen. Obviously, more small bone fragments were preserved in the midden deposits from the Wall site than in the pitfill at the Fredricks site. It should be noted that only those bones and bone fragments that appeared to be identifiable were pulled from the material recovered in the 1/16-inch screen. Thus, the percentage of small, identifiable fragments is actually higher in the Wall site assemblage than in the Fredricks site assemblage. At both sites, much of the bone recovered in the 1/2-inch screen consisted of identifiable fragments of bones of larger animals and complete, or nearly complete, bones from medium-sized animals. Identifiable bone from the 1/16-inch screen belonged, for the most part, to smaller species, such as fish and amphibians. The vast majority of the bone recovered in the 1/4-inch screen, however, consisted of fragments of bone that either were too small or too fragmented to be identified. The percentage of bone fragments that could not be identified was higher for the Fredricks site assemblage (47.28%) than for the Wall site assemblage (42.02%). It is likely that this is a result of the fact that the percentage of bone recovered in the 1/4-inch screen was also higher at the Fredricks site than at the Wall site.
Another way in which the condition of the bones from the two sites can be evaluated is by comparing the extent of fragmentation of the bones in the two assemblages. Extent of fragmentation can be determined from the number of fragments of deer bones present per individual identified (Runquist 1979:172). At the Wall site, a minimum of 36 individuals and 4,731 fragments were identified as white-tailed deer, which yields a ratio of 131.42 fragments per individual. For the Fredricks site, nine individuals and 1,128 fragments were identified as white-tailed deer, which yields a ratio of 125.33 fragments per individual. Thus it seems that, at least for the white-tailed deer, the bones in the Fredricks site assemblage are only slightly less fragmented than those in the Wall site assemblage. As noted earlier, the faunal remains from the Fredricks site may represent refuse cleaned from house floors, which would make the original contexts of the analyzed bone from both sites quite similar. Therefore, the slight difference in the ratios suggests to some extent that large bones deposited in pits may not be subjected to factors causing fragmentation as frequently as those deposited in sheet midden.
Almost 31% of the bone from the Fredricks site was burned whereas only 8.9% of the bone from the Wall site was burned. This suggests the possibility that the deposits from which the Fredricks site assemblage were derived represent a limited range of activities such as cleaning house floors or hearths. A greater range of activities that did not produce burned bone may be represented by the Wall site assemblage. Table 11 shows the percentage of deer skeletal elements represented in the Wall and Fredricks site assemblages. With the exception of six elements (innominate, atlas vertebrae, axis vertebrae, cervical vertebrae, sacrum, and patella) there is a higher percentage of every element represented at the Fredricks site than at the Wall site. This is one indication that the Fredricks site assemblage is better preserved than the Wall site assemblage. However, it could also be an indication that deer bones were treated differently by the inhabitants of the two sites. If, for example, the inhabitants of one of the sites frequently utilized deer bones as tools, it is possible that certain skeletal elements would not be discarded in the midden as food refuse. These tools would be curated, and thus would not be recovered in the midden in the same percentages as would be expected if preservation were the only factor being considered.
For example, proximal metacarpals and metatarsals should survive better than less-resistant elements such as proximal humeri or frontals. Proximal metacarpals and metatarsals were made into beamers by some Piedmont groups, and several of these hide-working tools have been identified in the Wall site assemblage (one in the 1983-1984 assemblage and nine in the assemblage examined by Runquist). As yet, however, no tools of this kind have been recovered from the Fredricks site, whose inhabitants had access to metal tools that may have made bone beamers obsolete. The percentages of proximal metacarpals (13.9%) and metatarsals (45.8%) recovered at the Wall site are not much higher than the percentages of other elements which could have been expected to be less well-preserved. At the Fredricks site, the percentages of proximal metacarpals (94.4%) and of metatarsals (77.8%) is considerably higher than the percentages for many of the other elements. Thus, it is likely that the different representation of deer skeletal elements at the two sites is a result of differential patterns of use and discard of the bones by the inhabitants of the two sites in addition to the possible effects of differential preservation.
There is no evidence, therefore, that the bone from one site is appreciably better preserved than the bone from the other site. It also follows that, in this case, that bones deposited in a pit are not better preserved than those discarded in an open midden. It is possible, however, that large bones deposited in pits will be slightly less fragmented than bones deposited in sheet midden.
Overall, the faunal assemblages from the Wall and Fredricks sites are very similar. Only seven species were identified at the Wall site that were not present in the Fredricks site assemblage. These were rabbit, beaver, chipmunk, muskrat, meadow vole, flying squirrel, and bluejay. With the exception of rabbit (MNI=4) and meadow vole (MNI=2), none of these species was represented by more than a single individual. Whereas meadow vole was represented by two individuals, it is very likely that these burrowing animals were intrusive in the deposit and were not utilized by the site's inhabitants. Rabbit is the only species from the Wall site assemblage that can be considered notable in its absence from the Fredricks site assemblage.
Ten species were identified from the Fredricks site that were not identified in the 1983-1984 assemblage from the Wall site. These were horse, pig, skunk, gray fox, mountain lion, red-bellied woodpecker, lesser scaup, musk turtle, sunfish, one individual belonging to the family Charadriidae (plover), and one individual belonging to the family Fringillidae (sparrow). As only one of these species, skunk, was present in the assemblage analyzed by Runquist, it is likely that none of these species was utilized to any great extent, if at all, by the inhabitants of the Wall site. With the exception of the sparrow (MNI=2), these species were only represented by a single individual each in the Fredricks site assemblage. The presence of two European-introduced mammals, pig and horse, in the Fredricks site assemblage is important. However, pig was represented by only one femur fragment and horse by only one molar.
Thus, based on the presence or absence of individual species, the data suggest there were no major differences in the utilization of faunal resources by the inhabitants of the two sites. The two exceptions noted are the absence of rabbit and the presence of two European domesticates in the Fredricks site assemblage.
Although the gross inventories of species utilized by the inhabitants of the two sites are virtually the same, there seem to be differences in the ways and relative amounts in which these species were procured and utilized. In an attempt to determine which species were most important in the diet of the inhabitants of the sites, the amount of meat available from each was calculated, using estimations by Smith (1975a), White (1953), and Cleland (1966). These figures are presented in Table 12. It should be noted that the bones, skins, furs, and carapaces of these animals were often important to the Indians as materials for tools, clothing, utensils, and other goods. Thus, a particular species would not always have been selected on the basis of its value as a source of food. The presence of only a single molar identified as horse indicates that this animal probably was not used for food by the inhabitants of the Fredricks site. Therefore, the amount of meat provided by this animal was not included in the calculations of available meat at this site.
The most important animals in the Wall site assemblage, listed in rank order of estimated meat yield, were deer, catfish, bear, raccoon, beaver, and turkey. At the Fredricks site the order was deer, bear, catfish, pig, mountain lion, turkey, and raccoon. Again, the assemblages appear to be quite similar.
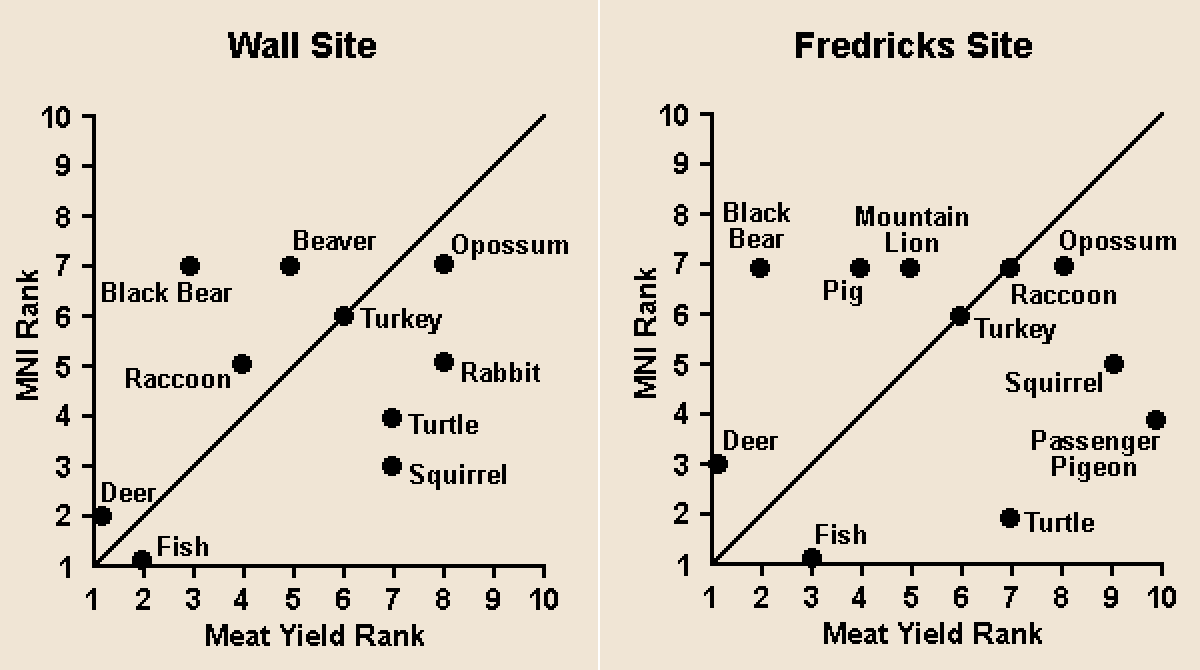
In an attempt to gain a more detailed indication of the relative importance of the various species utilized, 13 species or species groups were ranked according to a technique proposed by Smith (1975b:125-127). Using this approach, the species were ranked by their relative importance in terms of both the minimum number of individuals and projected meat yield (see bivariate plots).
At both sites, the species cluster into four groups. Deer and fish ranked very high on both scales and were evidently the most important faunal resources at the two sites. The second cluster consists of animals that ranked fairly high in terms of meat yield but were not frequently utilized. At the Wall site these animals were black bear, mountain lion, and beaver; at the Fredricks site, they were black bear and pig. Smith (1975b:126) notes that the low exploitation of bear and beaver at the Middle Mississippian sites whose faunal remains he analyzed may have been due to the fact that these species have low rates of reproduction, and thus were rarely encountered. It is interesting to note, however, that in his description of his visit to Occaneechi Town, Lawson (Lefler 1967:61) mentioned that the Indians brought him "good fat Bear" and that "Their Cabins were hung with a good sort of Tapestry, as fat Bear, and barbakued or dried Venison." It is also unlikely that pig would have been readily available to the inhabitants of the Fredricks site since this site was occupied early in the colonization process.
The third cluster consists of species that were utilized in high numbers but which yielded relatively small quantities of meat. At the Wall site, these species were rabbit, squirrel, and turtle. At the Fredricks site, they were turtle, squirrel, and passenger pigeon. At Fredricks, turtles were represented in higher numbers than were deer.
The fourth cluster of animals includes those species whose MNI and meat yield ranks were nearly equal. None of these species ranks highly in terms of either criterion of importance. At both sites these species were raccoon, turkey, and opossum.
At both sites, then, deer and catfish were the most important faunal resources. Turtle and squirrel were major secondary resources, as was rabbit at the Wall site and passenger pigeon at the Fredricks site. Raccoon, turkey, and opossum were utilized on a more limited basis at both sites. Bear and beaver at the Wall site, and pig and mountain lion at the Fredricks site, provided large quantities of meat but were not as frequently encountered as were other species.
{kind=link}